


Alte chemische Stoffe, die Carotinoide, sind allgegenwärtig und dienen einer Reihe von Zwecken. Als Membranstabilisatoren und UV-Schutzmittel kamen diese Verbindungen ursprünglich in der Welt der Mikroben, insbesondere in Archaeen, vor.
Das lineare Polytetraen (C40), von dem sich fast alle Carotinoide ableiten, weist Doppelbindungen auf, die diese Moleküle dazu veranlassen, Licht zu absorbieren. Dies ist eine Folge des Überflusses an Carotinoiden in entwickelten Organismen und ihrer Verfügbarkeit für die Entwicklung neuartiger Aktivitäten. Die Entstehung bestimmter Botenmoleküle aus Carotinoid-Vorläufern und ihren sauerstoffhaltigen Derivaten ist ein Beispiel dafür, dass eine Gruppe von Chemikalien in entwickelten Organismen viele Funktionen hat.
Die oxidative Umwandlung von Carotinoiden in spezifische Produkte des regulatorischen Abbaus von Apocarotenoiden, die von einer Multi-Enzym-Familie von Carotinoid-abbauenden Dioxygenasen (CCDs) katalysiert wird, ist ein weiterer Ausdruck der Evolution von Bakterien zu Säugetieren.
Die Geschwindigkeit, mit der der Carotinoidweg das Substrat liefern kann, begrenzt die Geschwindigkeit, mit der Apocarotenoide gebildet werden können, was im Wesentlichen ein erweiterter Weg für Carotinoide ist. Zwei wichtige Carotinoide, die als Substrate für die Bildung der charakteristischen Apocarotenoide des Safrans dienen, sind Zeaxanthin und Beta-Carotin.
Carotinoide in Plastiden verbinden die lineare Säule von Cyclase- und Lyquin-Cyclase-Vorläufern wieder zu terminalen Ionenringen. Die Strukturen dieser Ringe sind jedoch voneinander verschieden. Durch Anhängen eines Betarings an Lycopin durch die Lycopin-Beta-Cyclase entsteht Gamma-Carotin, und das Anhängen eines weiteren Betarings an sein freies Ende durch dasselbe Enzym führt zur Bildung des orangen Betacarotin-Pigments. Die Hydroxylierung von zwei Beta-Ringen in Beta-Carotin während einer Reaktion, die von der Beta-Carotin-Hydroxylase (BCH) katalysiert wird, führt zur Bildung von Zeaxanthin. Sowohl Beta-Carotin als auch Zeaxanthin dienen als Vorstufen für die Biosynthese der einzigartigen Apocarotenoide des Safrans.
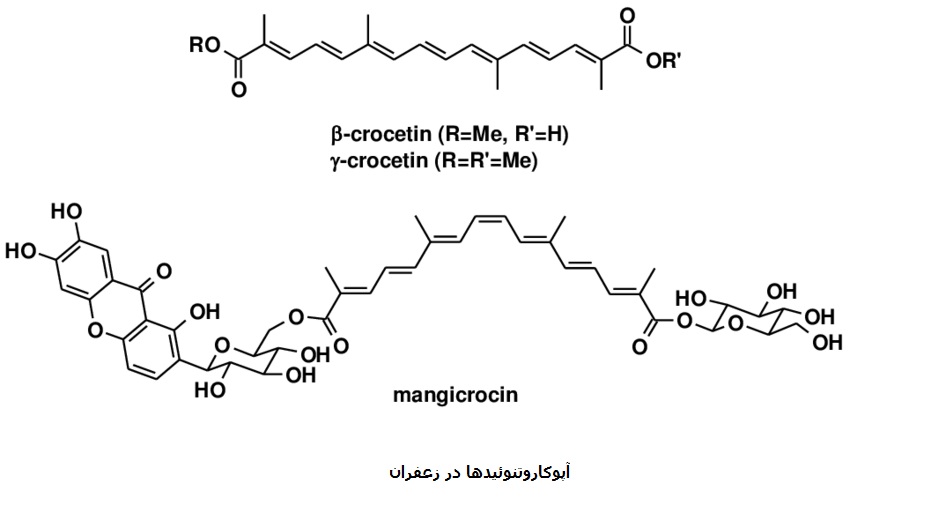
Safran und Carotinoide
Wiederholte strukturelle Segmente in der Vielzahl der bekannten Apocarotenoide deuten auf die Möglichkeit hin, dass diese Segmente durch typische biosynthetische Prozesse aus Carotinoiden in voller Länge entstanden sind.
Zeaxanthin und Beta-Carotin sowie Carotinoid-abbauende Dioxygenasen (CCD) werden in Safran zu Apocarotenoiden abgebaut. Typischerweise ist diese Familie durch eine amphipathische alpha-helikale Sequenz gekennzeichnet. Vier konservierte Histidinreste in dieser katalytischen Region sind für die dative Bindung eines Peptids als Permeant und von Nichteisen-Eisen (Fe) als Cofaktor durch die Plastidenmembran notwendig.
Einige Carotinoid-abbauende Dioxygenasen, die im Zytoplasma angesiedelt sind (z. B. (CCD)), bilden eine Ausnahme von dieser Regel. Man geht davon aus, dass Carotinoid-abbauende Dioxygenasen mit zwei pflanzlichen oder tierischen Ursprüngen die gleiche Funktion haben, da es sich bei beiden um Atome handelt. Der zum Sauerstoffmolekül gehörende Sauerstoff ist über eine Doppelbindung mit seinem Substrat verbunden und bewirkt die Bildung eines aldehydhaltigen Produkts.
Es war immer umstritten, ob es sich bei diesen Enzymen um eine Monooxygenase oder eine Dioxygenase handelt. Mit Hilfe von Markierungstests konnte jedoch eindeutig nachgewiesen werden, dass diese Enzyme über einen Dioxygenase-Mechanismus verfügen. Für die Apocarotenoid 15, 15-prime Oxygenase (ACO), ein Enzym aus dem Cyanobakterium Cinchocystis, wurde gerade seine Kristallstruktur effektiv charakterisiert. Es wurde festgestellt, dass dieses Enzym eine Typ-III-Struktur in Form eines Beta-Schmetterlings mit sieben Flügeln aufweist.
Die Kristallstruktur des ACO-Proteins hat gezeigt, dass das aktive Zentrum des Enzyms vier abgeschirmte Histidinreste und ein Eisenion enthält. Alle Mitglieder der Multienzymfamilie der Carotinoid-abbauenden Dioxygenasen haben diese Anordnung von vier Eisenhistidinen gemeinsam, wenngleich sich ihre Aminosäuresequenzen ansonsten leicht unterscheiden. Darüber hinaus wurde festgestellt, dass drei aufeinanderfolgende Doppelbindungen während des katalytischen Prozesses von der reinen trans-Form in eine gebogene cis-trans-cis-Struktur umgewandelt werden, wobei die letzte trans-Bindung in Fe2, das an Dioxygen gekoppelt ist, abgelagert und durch Sauerstoff abgebaut wird.
In Bezug auf die Gesamtsequenz, den Schutz von Histidinabschnitten, die Verwendung von Eisen als Cofaktor und die Fähigkeit, Doppelbindungen zu spalten, sind sich die Carotinoid-abbauenden Oxygenasen in Pflanzen und die Lignostylendioxygenasen in Bakterien bemerkenswert ähnlich, was darauf hindeutet, dass die beiden Enzyme von einem gemeinsamen Vorfahren abstammen könnten. Jüngste Forschungen an einigen Dioxygenasen, die Carotinoide abbauen, haben gezeigt, dass RPE65, NinaB und die meisten Dioxygenasen, die Carotinoide in Säugetieren abbauen, eine inhärente Isomerase-Aktivität aufweisen. Bei den Dioxygenasen, die Carotinoide in Pflanzen abbauen, wurde jedoch keine inhärente Isomeraseaktivität festgestellt, was darauf zurückzuführen sein könnte, dass eisenabhängige Carotinoid-Isomerasen separat existieren.
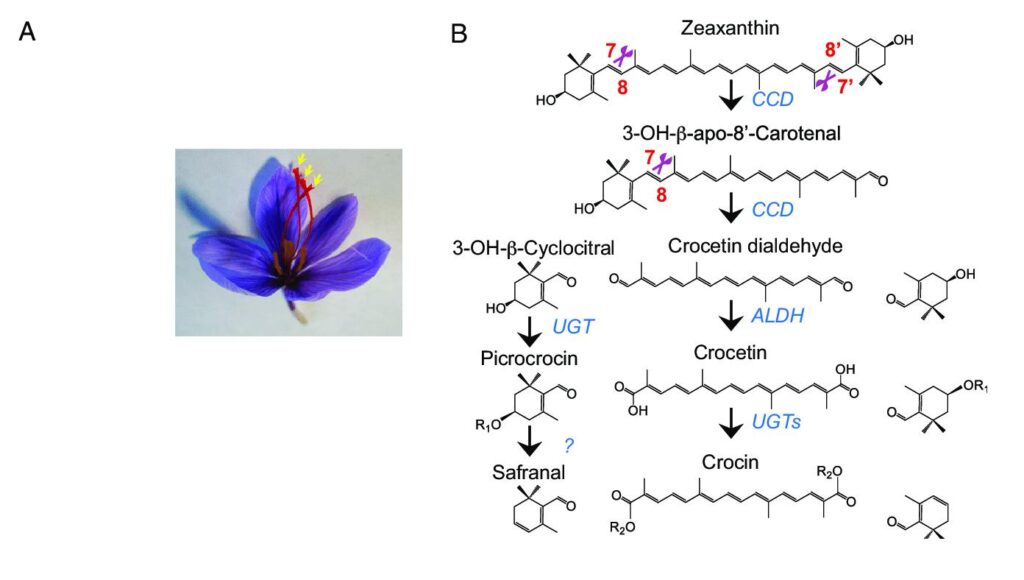
In Safran waren die ersten Carotinoid-abbauenden Dioxygenasen in der Biosynthese von Apocarotenoiden, die kloniert, identifiziert und funktionell bestimmt wurden, die Xenaxanthin-abbauende Dioxygenase (CSZCD) und die Carotinoid-abbauende Dioxygenase (9-prime und 10-prime). Es wurde festgestellt, dass CSZCD nur im Rahmgewebe exprimiert wird und unter Dehydratationsstress zunimmt, während die Expression von CSCCD durch Dehydratationsstress nicht beeinträchtigt wird und im Blüten- und Blattgewebe konstant ist.
Die Ergebnisse der Untersuchungen zeigen einen sequenziellen Prozess, der mit dem oxidativen Abbau von Zeaxanthin innerhalb der Chromoplasten beginnt und mit der Ablagerung der wasserlöslichen Nebenprodukte in der zentralen Vakuole endet. Zwei Isoformen von CSCCD und drei Isoformen von CSCCD4 wurden kürzlich auch in Safran entdeckt.
Trotz des Expressionsmusters von CSCCDfa wurde auch festgestellt, dass CSCCDa eine einheitliche Expression aufweist und CSCCDb sich ausschließlich in der Narbe manifestiert. Die -Ionen-Emission von -Carotin während der Narbenbildung wird durch CSCCFb und CSCCDFC dargestellt. Die Überexpression von CSCCD-Isoformen in bakteriellen Spezies, die Beta-Carotin akkumulieren, lieferte den Beweis für ihre Rolle bei der Erzeugung von Beta-Ionen. Andere Pflanzen, darunter die Nipapalme und die Tomate sowie Safran, verfügen über Carotinoid abbauende Dioxygenasen, die die Freisetzung von Beta- und Alpha-Ionen regulieren.
Beta-Ionon aus der Familie der Apocarotine spielt in Pflanzen eine wichtige Rolle; während Ionon vor allem Käfer (Anomala transvaalensis) abwehrt, ist Alpha-Ionon wirksamer gegen blütenfressende Schädlinge, die Pflanzen ernsthaft schädigen können. Im Vergleich zu den anderen war die Menge an Beta-Ionon aus Cheddar-Saft geringer. Es wurde festgestellt, dass transgene Arabidopsis mit vorexprimiertem CCD1 einen höheren Gehalt an Alpha-Ionon aufweist. Die Wirkung des Enzyms CCD-Dioxygenase führte zur Produktion dieses Apocarotenoids. Darüber hinaus wurde festgestellt, dass der Flohkäfer Phyllotreta cruciferae die transgenen Pflanzen weniger stark schädigt als die nicht-transgenen Wildpflanzen, wenn die transgenen Pflanzen mit einem erhöhten Alpha-Ionen-Gehalt gegen ihn getestet wurden.
Damit wurde nachgewiesen, dass diese Pflanzen durch Carotinoide vor Insektenfraß geschützt sind. Diese Stoffe dienen auch dem männlichen Gametophyten zur Abwehr von Bestäubern, die für die Pflanze schädlich sind.
Kürzlich wurde aus der Safran-Narbe CCD2 extrahiert, eine neue Carotinoid-abbauende Oxygenase. Im Gegensatz zu CSCCD und CSCCdf hat die Isoform CSCCDY eine andere Wirkungsweise. Diese Isoform und CSCCD sind nach phylogenetischen Analysen eng miteinander verwandt. Safran wurde auch mit CSCCDY und CSCCDA in Verbindung gebracht. Die Analysen der Sequenz und der phylogenetischen Themen der CSCCDY- und CSCCDA-Gene haben gezeigt, dass ihre evolutionären Prozesse vergleichbar sind, obwohl eine solche Ähnlichkeit in der CCD- und CCD-Unterfamilie nicht entdeckt wurde.
Die Paarung von 42 CCDs führte schließlich zur Entstehung von zwei Linien, die sich wiederum zu CCDY und CCDA entwickelten, wie die phylogenetischen Analysen von CSCCDY und CSCCDA zeigen. Es wird exprimiert, und die Entfernung der Knospenspitze reduziert seine Expression dramatisch, was zeigt, dass es an der Hemmung der lateralen Knospenexpansion beteiligt ist. Im Gegensatz zum vaskulären Gewebe der apikalen Knospe, wo die CSCCDA-Isoform in hohem Maße exprimiert wird, ist die CSCCDY-Isoform in der neu gebildeten Vaskulatur der lateralen Knospen stark ausgeprägt.
Während CCDY- und CCDA-Substrate in Safran nicht identifiziert wurden, führen diese beiden Enzyme in anderen Pflanzen durch den schrittweisen Abbau von Beta-Carotin schließlich zur Produktion von Strigolacton und einer neuen Art von Apocarotenoid-Hormon, das an der Stammverzweigung beteiligt ist.
Strigolactone wurden zunächst in der Substanz entdeckt, die aus der Wurzel der parasitären Pflanze Ajozeh-Gras abgesondert wird. Diese Verbindungen werden in Pflanzenwurzeln unter Bedingungen der Nährstoffbegrenzung gebildet und fördern das Wachstum von Seitenwurzeln und Haarwurzeln und verhindern das Wachstum von Seitenästen und Trieben der oberirdischen Pflanzenteile.
Durch diese Taktik wird die Anforderung von Nahrungsmitteln durch den Spross reduziert und gleichzeitig die Aufnahme von Nährstoffen durch die Wurzeln erhöht. Die jetzt vorliegenden Erkenntnisse deuten darauf hin, dass Strychnolactone nur in Kombination mit anderen Phytohormonen wirksam sind, um das Wachstum der Bodenstängel (Stolen) zu kontrollieren und die Knollenruhe in dieser Pflanze aufrechtzuerhalten. Die Wirkungen dieser Verbindungen wurden durch Untersuchungen an der CCDA-RNA von Kartoffeln bestätigt, die mehr Haupt- und Nebenzweige als Kontrollpflanzen und eine geringere Ausläuferbildung sowie Zwergwuchs und eine fehlende Blüte in den Linien aufwiesen, die stark von diesen Phytohormonen beeinflusst wurden.
Mit seinem Angebot an natürlichen und nachhaltige Produkten versucht BioElixiro, das auf eine lange Geschichte der Erforschung, Prüfung und Herstellung biologischer Materialien zurückblicken kann, einen wichtigen Beitrag zur Gesundheit seiner Kunden zu leisten.Die folgenden Bio- und Naturprodukte sind bei der Golden Mart Business Group (BioElixiro) erhältlich und werden aus hochwertigen Rohstoffen hergestellt.
– Nüsse (wie Cashews, Haselnüsse, Mandeln, Rosinen und Pistazien).
• Trockenfrüchten
• Gewürze (Safran, Kardamom, Kümmel usw.)
• Berberitze
• Safran und und ihre Derivate, darunter:
• Safransorten (super negin, negin, sargol, all red, Usw .)
• SafranSirup
• Safran-Kandiszucker
• Tee und Kräutertee
Bestellen Sie einfach das Produkt BioElixiro bei den Spezialisten von Golden Mart, um diese Artikel überall auf der Welt zu erhalten.