

 طلا و جواهرات
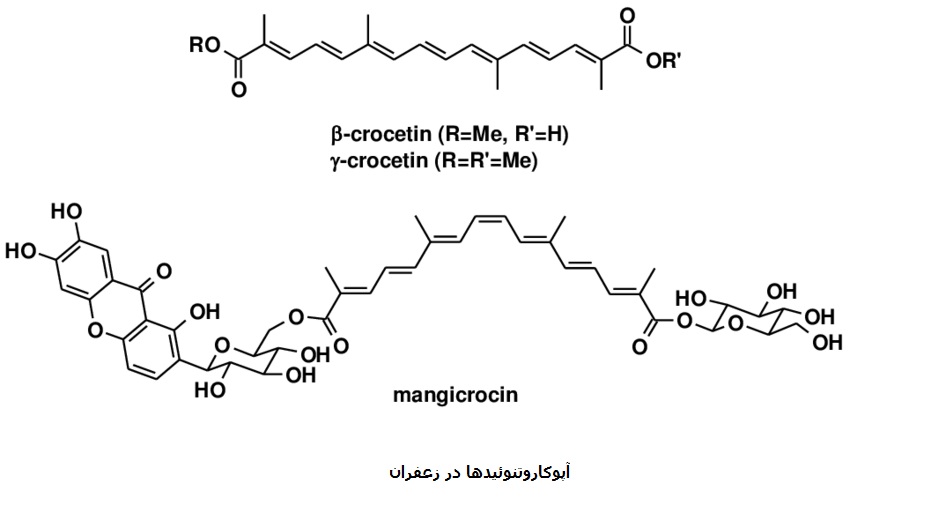
طلا و جواهراتکاروتنوئیدها مولکلولهایی با قدمتی دیرینه اند که به وفور یافت شده و عملکردهای مختلفی دارند این مولکلولها در ابتدا در دنیای میکروبی از جمله در آرکایا بعنوان تثبیت کننده های غشایی و محافظت کننده در مقابل اشعه ماوراء بنفش پدیدار شدند.
تقریبا تمامی کاروتنوئیدها از یک پلین تتراترین خطی (C۴۰) بوجود می آیند که حضور باندهای دوگانه مضاعف سبب جذب نور توسط این ترکیبات می.شود فراوانی کاروتنوئیدها در موجودات تکامل یافته و در نتیجه در دسترس بودن آنها برای توسعه کارکردهای جدید باعث شده است تا این دسته از ترکیبات کارکردهای متنوعی در موجودات تکامل یافته داشته باشند که بعنوان مثال میتوان به تکامل برخی مولکولهای پیام رسان از پیش سازهای کاروتنوئیدی و مشتقات اکسیژنه آنها اشاره نمود.
تبدیل اکسیداتیو کاروتنوئیدها به محصولات خاص تجزیه تنظیمی آپوکاروتنوئیدها که توسط خانواده ای چند آنزیمی از دی اکسیژنازهای حفظ ساختار شده تجزیه کننده کاروتنوئید (CCDs) کاتالیز میشود بیان دیگری از سیر تکامل از باکتریها به پستانداران است .
بیوسنتز آپوکاروتنوئیدها در واقع همان مسیر بسط یافته بیوسنتز کاروتنوئیدهاست و بواسطه سرعت فراهم شدن سوبسترا بوسیله مسیر کاروتنوئیدها محدود می.شود زئازانتین و بتا کاروتن دو کاروتنوئید مهم هستند که بعنوان سوبسترای بیوسنتز آپوکاروتنوئیدهای منحصر به فرد زعفران عمل مینمایند .
کاروتنوئیدها در پلاستیدها مجددا از پیش سازهای سیکلاز و هم لیکوین سیکلاز ستون خطی را بهم وصل مینمایند تا حلقههای یونون انتهایی را بوجود آورند .هرچند ساختارهای این حلقه ها از یکدیر متمایز هستند. افزودن یک حلقه بتا به لیکوپن بوسیله لیکوپن بتا سیکلاز باعث ایجاد گاما کاروتن میشود و افزودن یک حلقه بتای دیگر به انتهای آزاد آن توسط همین آنزیم منجر به تولید رنگیزه بتاکاروتن نارنجی خواهد شد. هیدروکسیلاسیون دو حلقه بتا در بتا کاروتن طی واکنشی که توسط بتا کاروتن هیدروکسیلاز (BCH) کاتالیز میشود باعث بوجود آمدن زئازانتین خواهد شد. هم بتا کاروتن و هم زئازانتین بعنوان پیش ماده بیوسنتز آپوکاروتنوئیدهای منحصر به فرد زعفران عمل می نمایند.
زعفران و کاروتنوئیدها
بخشهای ساختاری تکرار شونده که در طیف متنوع آپوکاروتنوئیدهای شناخته شده یافت شده اند این نکته را خاطر نشان می کند که این بخش ها از کاروتنوئیدهایی با طول کامل و احتمالا در مسیرهای بیوسنتزی ،معمول مشتق شده اند .
در زعفران، آپوکاروتنوئیدها بواسطه تجزیه کاروتنوئیدها و زئازانتین و بتا کاروتن و توسط دی اکسیژنازهای تجزیه کننده کاروتنوئید (CCD) بوجود می آیند . این خانواده عموما با یک دنباله مارپیچ آلفای آمفیپاتیک تشخیص داده میشود این دنباله یک دنباله کاتالیتیک است که در بر گیرنده چهار بخش هیستیدینی محافظت شده مسئول پیوند داتیو آهن غیر هم آهن فروس (Fe) بعنوان ،کوفاکتور و یک پپتید بعنوان مجوز عبور از غشاء پلاستیدی میباشد.
برخی دی اکسیژنازهای تجزیه کننده کاروتنوئید که در سیتوپلاسم قرار گرفته اند (مانند (CCD) از این قاعده مستثنی هستند. تصور بر این است که دی اکسیژنازهای تجزیه کننده کاروتنوئید با دو منشا گیاهی یا ،جانوری دارای عملکرد یکسانی هستند؛ زیرا هر دوی آنها اتمهای اکسیژن متعلق به مولکول اکسیژن را توسط یک پیوند دوگانه به سوبسترای خود متصل نموده و سبب ایجاد محصول حاوی آلدهید می شوند.
همواره بر سر مونواکسیژناز یا دی اکسیژناز بودن مکانیزم این آنزیم ها اختلاف وجود داشته است؛ هرچند شواهد قانع کننده ای مبنی بر مکانیزم دی اکسیژناز این آنزیم ها وجود دارد که از آزمایشات برچسب دار بدست آمده است . اخیرا ساختار کریستالی آپو کاروتنوئید ۱۵، ۱۵ پریم اکسیژناز (ACO) متعلق به سیانوباکتریوم سینکوسیستیس با موفقیت تعیین و مشخص شد که این آنزیم دارای ساختار نوع سوم بصورت پروانه ای بتا با ۷ تیغه است .
ساختار کریستالی پروتئین ACO نشان داده است که آنزیم دارای یک یون Fer پیوند شده به چهار بخش هیستیدینی محافظت شده در سایت فعال است. این آرایش چهار هیستیدین آهن در بین تمامی اعضاء خانواده چند آنزیمی دی اکسیژنازهای تجزیه کننده کاروتنوئید مشترک است هرچند غیر از این یک ویژگی توالی های اسیدهای آمینه آنها تا حدودی ناهمسان است. بعلاوه گزارش شده است که در طی فرآیند کاتالیز سه باند دوگانه متوالی از فرم تمام ترانس به یک ساختار خمیده سیس ترانس سیس تبدیل میشود که باند ترانس باقیمانده در Fe۲ متصل به دی اکسیژن قرار گرفته و توسط اکسیژن تجزیه میشود .
اکسیژنازهای تجزیه کننده کاروتنوئیدها در گیاهان از نظر توالی کلی، حفاظت بخش های هیستیدینی، نقش آهن به عنوان کوفاکتور و شکست باندهای دوگانه شباهت زیادی به لیگنوستیلین دی اکسیژنازها (LSD) ی باکتریها دارند که نشان میدهد آنزیم ها ممکن است از یک نیای مشترک تکامل یافته باشند .مطالعات اخیر روی برخی دی اکسیژناز های تجزیه کننده کاروتنوئید ها نشان دهنده وجود فعالیت ایزومراز ذاتی از جمله در RPE۶۵ و NinaB و بطور کلی دی اکسیژنازهای تجزیه کننده کاروتنوئیدهای پستانداران است .هر چند هیچگونه فعالیت ایزومراز ذاتی در دی اکسیژناز های تجزیه کننده کاروتنوئیدها در گیاهان مشاهده نشده است که میتواند بدلیل حضور مستقل ایزومرازهای کاروتنوئیدی وابسته به آهن باشد .
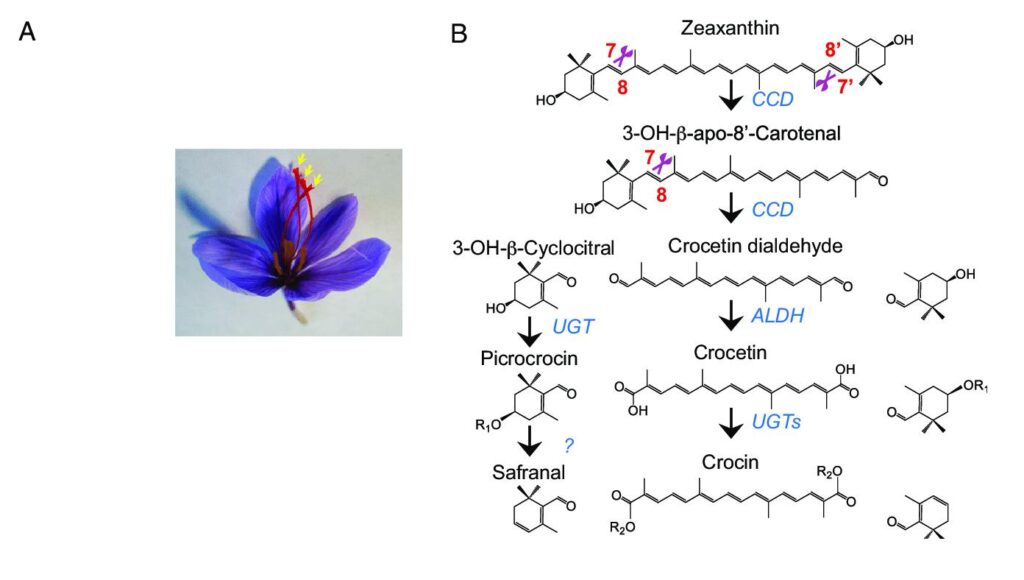
در زعفران اولین دی اکسیژنازهای تجزیه کننده کاروتنوئید در بیوسنتز آپوکاروتنوئیدها که کلون سازی، شناسایی و بلحاظ کارکردی مشخص شدند دی اکسیژناز تجزیه کننده زنازانتین (CSZCD) و دی اکسیژناز تجزیه کننده کاروتنوئید(9 پریم 10 پریم) بودند.مشخص شده است که CSZCD تنها در بافت خامه بیان میشود و تحت تنش کم آبی نیز افزایش مییابد در حالیکه بیان CSCCD بدون تاثیر پذیری از تنش کم آبی و با سرعت ثابت در بافت گل و برگ انجام می گیرد.
مطالعات صورت گرفته وجود نوعی توالی گام به گام را نشان میدهد که شامل تجزیه اکسیداتیو زئازانتین داخل کروموپلاستها و سپس ترسیب محصولات بدست آمده از تجزیه این محصولات قابل حل در آب هستند در واکوئل مرکزی میشود . اخیرا دو ایزوفرم CSCCD و سه ایزوفرم CSCCD۴ نیز در زعفران شناسایی شدند که علاوه بر مشخص شدن کارکرد آنها معلوم شد که در بیوسنتز بتا یونون هم دخالت دارند.
همچنین مشاهده شد که بیان CSCCDa بصورت یکنواخت بوده و CSCCDb نیز تنها در کلاله بیان می شود ،هر چند الگوی بیان CSCCDfa. CSCCFb و CSCCDFC با انتشار بتا یونون بدست آمده از بتا کاروتن در طی نمو کلاله مطابقت دارد. عملکرد ایزوفرمهای CSCCD تولید بتا یونون توسط بیش بیان آنها در گونه های باکتریایی که تجمع بتا کاروتن در آنها صورت می گیرد تایید شد . علاوه بر زعفران دی اکسیژنازهای تجزیه کننده کاروتنوئید همچنین انتشار بتا و آلفا یونون در دیگر گیاهان مانند نخل نیپا و گوجه فرنگی را نیز کنترل میکنند .
آپو کاروتنوئید بتا یونون در گیاهان وظایف مهمی بر عهده دارد در حالیکه بنا یونون بطور اختصاصی سبب دفع سوسک ها (Anomala transvaalensis می شود، آلفا یونون بیشتر در برابر آفت گل خورم میشود که میتواند سبب بروز آسیب جدی به گیاهان شود . تغذیه نوعی کنه زمین از شیره شیدر بدلیل داشتن سطح بالایی از بتا یونون در مقایسه با دیگر ترینها کمتر بود .انجام گرفت مشخص گردید که پیش بیان CCD۱ در آرابیدوپسیس تراریخته باعث افزایش سطح آلفا یونون شد. این آپو کاروتنوئید قرار در نتیجه فعالیت دی اکسیژناز CCD تولید شده شده بود. بعلاوه وقتیکه گیاهان تراریخته که سطح آلفا یونون در آنها افزایش یافته بود در مواجهه با سوسک کک مانند چلیپائیان ۴۱ Phyllotreta cruciferae مورد آزمایش قرار گرفتند معلوم شد که خسارت ناشی از تغذیه این سوسک در گیاهان تراریخته در مقایسه با گیاهان تیب وحشی غیر تراریخته کاهش یافته است .
این موضوع نشان داد که کاروتنوئیدهای قرار سبب جلوگیری از تغذیه حشرات از این گیاهان میشوند. این ترکیبات همچنین منجر به حفاظت از گامتوفیت نر در مقابل موجودات گرده خواری شد که برای گیاه منفعتی ندارند
اخیرا نوع جدیدی از اکسیژنازهای تجزیه کننده کاروتنوئید بنام CCD۲ از کلاله زعفران جداسازی گردید .نوع فعالیت ایزوفرم CSCCDY با CSCCD و CSCCdf با هم متفاوت است تجزیه و تحلیلهای فیلوژنتیکی نشان داده اند که این ایزوفرم ارتباط نزدیکی با CSCCD دارد CSCCDY و CSCCDA نیز در زعفران شناسایی شده اند تجزیه و تحلیلهای صورت گرفته روی توالی و مباحث فیلوژنتیکی CSCCDY و CSCCDA مشخص نموده است که روند تکاملی ژن های CCDY CCDA به هم شباهت دارند؛ در حالیکه چنین شباهتی در زیر خانواده CCD و CCD یافت نشده است.
بعلاوه تجزیه و تحلیلهای فیلوژنتیکی CSCCDY و CSCCDA سرنخی در ارتباط با همتاسازی ۴۲ CCD ارائه می دهد که نهایتا منجر به ظهور دو تبار میشود و این دو به CCDY و CCDA تکامل می.یابند در ،زعفران CSCCDA در جوانه های جانبی در حال سکون به شدت بیان می شود و بیان آن بطور معنی داری با بریدن راس جوانه کاهش می یابد که نشان میدهد در سرکوب رشد جوانه جانبی دخالت دارد. برخلاف CSCCDA ایزوفرم CSCCDY در یافت آوندی تازه توسعه یافته جوانه های جانبی در مقایسه با بافت آوندی جوانه ،راسی به شدت بیان میشود .
در حالیکه سوبستراهای CCDY و CCDA در زعفران مشخص نشده اند در دیگر گیاهان این دو آنزیم با تجزیه گام به گام بتا کاروتن در نهایت منجر به تولید استریگولاکتون و نوعی هورمون آپوکاروتنوئیدی جدید می شوند که در شاخه زایی ساقه دخالت دارد .
استریگولاکتون ها، در ابتدا در ماده مترشحه از ریشه گیاه انگلی علف عجوزه کشف شدند. این ترکیبات در شرایط محدودیت مواد غذایی در ریشه های گیاه تولید و سبب افزایش رشد ریشه های جانبی و ریشه های مویین شده و از رشد شاخه های جانبی و جوانههای اندام هوایی گیاه جلوگیری می کند .
این استراتژی سبب افزایش جذب مواد غذایی توسط ریشه شده و همزمان درخواست اندام هوایی برای دستیابی به منابع غذایی را محدود می کند. بر اساس گزارشات موجود استرایکولاکتونها بطور انحصاری با به همراه سایر فیتوهورمونها در مورفولوژی سیب زمینی و همچنین هدایت رشد ساقه های زمینی (استولن) و حفظ خواب غده ها در این گیاه موثر است. مطالعاتی که بر روی CCDA RNA سیب زمینیهایی که تعداد شاخه های فرعی و اصلی آنها در مقایسه با بوتههای شاهد بیشتر و تولید استولون در آنها کمتر بود در کنار ایجاد کوتولگی و عدم گلدهی در لاینهایی که به شدت تحت تاثیر این فیتوهورمونها قرار گرفته بودند اثرات این ترکیبات در سیب زمینی را تایید نمودند .
بیو الکسیرو (Bio Elixiro)با سابقه طولانی در تحقیق ،آزمایش و تولید مواد ارگانیک و داشتن چندین شعبه در سراسر دنیا تلاش میکند ،تا با ارائه محصولات طبیعی و ارگانیک گامی بلند برای سلامتی مشتریان خود بردارد.گروه تجاری گلدن مارت تحت برند بیو الکسیرو (Bio Elixiro) محصولات ارگانیک و طبیعی زیر را با مواد اولیه با کیفیت و بسیار مرغوب ارائه میدهد.
• -خشکبار (پسته ، بادام،کشمش ،فندق ،بادام هندی و…)
• -میوه خشک
• -ادویه جات (زعفران ،هل ،زیره و…)
• -زرشک
• -زعفران و مشتقات آن از قبیل:
• انواع زعفران (سوپر نگین ،نگین ،سرگل ،دخترپیچ و…)
• شربت زعفران
• نبات زعفرانی
• چای و دمنوش
برای تهیه این محصولات در هرجایی از دنیا که هستید فقط کافی است با کارشناسان گلدن مارت تماس بگیرید و محصول بیو الکسیرو (Bio Elixiro) را سفارش دهید.